

Что такое электромеханическое сопряжение. Феномен электромеханического сопряжения
Электромеханическое сопряжение в мышцах
Электромеханическое сопряжение – это цикл последовательных процессов, начинающийся с возникновения потенциала действия ПД на сарколемме (клеточной мембране) и заканчивающийся сократительным ответом мышцы.
Основная особенность электронных учебных пособий состоит в сочетании необходимого и специально подобранного теоретического материала, дополняющего печатные издания с большим числом разнообразных, тщательно проработанных тестов.
Многофункциональное электронное учебное пособие предназначено для самостоятельного освоения учебного курса, получения навыков практического применения знаний, для автоматизации и интеллектуализации прикладных задач. Ориентация системы тестирования на личностные качества обучаемых позволяет определить индивидуальные особенности обучающегося и, в соответствии с этим, рекомендовать методику обучения, которая позволит оптимизировать процесс получения знаний.
Нарушение последовательности процессов сопряжения может приводить к патологиям и даже к летальному исходу. Основные этапы этого процесса можно проследить по схеме рисунок 11.

Рисунок 11 Схема электромеханического сопряжения в кардиомиоците (М – клеточная мембрана-сарколемма, СР – саркоплазматический ретикулум, МФ – миофибрилла, Z – z-диски, Т – Т-система поперечных трубочек); 1 – поступления Na+ и 2 – поступления Са2+ в клетку при возбуждении мембраны, 3 – “кальциевый залп”, 4 – активный транспорт Са2+ в СР, 5 – выход из клетки К+, вызывающий реполяризацию мембраны, 6 — активный транспорт Са2+ из клетки
Процесс сокращения кардиомиоцита происходит следующим образом (номера пунктов в тексте соответствуют номерам процессов в схеме электромеханического сопряжения на рисунок 11):
- 1 – при подаче на клетку стимулирующего импульса открываются быстрые (время активации 2 мс) натриевые каналы, ионы Na+ входят в клетку, вызывая деполяризацию мембраны;
- 2 – в результате деполяризация плазматической мембраны в ней и в Т-трубочках открываются потенциал-зависимые медленные кальциевые каналы (время жизни 200 мс), и ионы Са2+ поступают из внеклеточной среды, где их концентрация 2 * 10-3 моль / л, внутрь клетки (внутриклеточная концентрация Са2+ 10-7 моль /л);
- 3 – кальций, поступающий в клетку, активирует мембрану СР, являющегося внутриклеточным депо ионов Са2+ (в СР их концентрация достигает = 10″3 моль/л), и высвобождает кальций из пузырьков СР, в результате чего возникает так называемый «кальциевый залп». Ионы Са2+ из СР поступают на актин-миозиновый комплекс МФ, открывают активные центры актиновых цепей, вызывая замыкание мостиков и дальнейшее развитие силы и укорочения саркомера;
- 4 – по окончании процесса сокращения миофибрилл ионы Са2+ с помощью кальциевых насосов, находящихся в мембране СР, активно заканчиваются внутрь саркоплазматического ретикулума;
- 5 – процесс электромеханического сопряжения заканчивается тем, что К+ пассивно выходит из клетки, вызывая реполяризацию мембраны;
- 6 – ионы Са2+ активно выводятся во внеклеточную среду с помощью кальциевых насосов сарколеммы.
Таким образом, в кардиомиоците электромеханическое сопряжение идет в две ступени: вначале небольшой входящий поток кальция активирует мембраны СР, способствуя большему выбросу кальция из внутриклеточного депо, а затем в результате этого выброса происходит сокращение саркомера. Описанный выше двухступенчатый процесс сопряжения доказан экспериментально. Опыты показали, что: а) отсутствие потока кальция извне клетки jCa прекращает сокращение саркомеров, б) в условиях постоянства количества кальция, высвобождаемого из СР, изменение амплитуды потока кальция приводит к хорошо коррелирующему изменению силы сокращения.
Следует отметить, что не во всех мышечных клетках организма процесс сопряжения происходит, как в кардиомиоците. Так, в скелетных мышцах теплокровных потенциал действия короткий (2-3 мс) и медленный поток ионов кальция в них отсутствует. В этих клетках сильно развита Т-система поперечных трубочек, подходящих непосредственно к саркомерам близко к z-дискам (см. рисунок 11). Изменения мембранного потенциала во время деполяризации через Т-систему, вызывая залповое высвобождение ионов Са2+ и дальнейшую активацию сокращения (3, 4, 5).
Временной ход описанных процессов показан на рисунок 12.
Общим для любых мышечных клеток является процесс освобождения ионов Са2+ и внутриклеточных депо – саркоплазматического ретикулума и дальнейшая активация сокращения. Ход кальциевого выброса из СР экспериментально наблюдается с помощью люминесцирующего в присунокутствии ионов Са2+ белка экворина, который был выделен из светящихся медуз. Задержка начала развития сокращения в скелетных мышцах составляет 20 мс, а в сердечной – несколько больше (до 100 мс).

Рисунок 12 Временное соотношение между потенциалом действия кардиомиоцита (а) и одиночным сокращением (б) в этих клетках. Ордината слева – мембранный потенциал, справа – сила. – потенциал покоя
Феномен электромеханического сопряжения
– соотношение между временным ходом потенциала действия в мышечном волокне и возникающим в результате этого сокращением мышечного волокна с последующим его расслаблением.
Это последовательность процессов, в результате которых потенциал действия плазматической мембраны мышечного волокна приводит к запуску сокращения мышцы или к так называемому циклу поперечных мостиков, который будет продемонстрирован далее.
Плазматическая мембрана скелетных мышц электрически возбудима и способна генерировать распространяющийся потенциал действия посредством механизма, аналогичного тому, который действует в нервных клетках. Потенциал действия в волокне скелетной мышцы длится 1-2 мс и заканчивается раньше, чем появятся какие-либо признаки механической активности (рис. 12). Начавшаяся механическая активность может продолжаться более 100 мс. Электрическая активность плазматической мембраны не оказывает прямоговлияния на сократительные белки, а вызывает повышение цитоплазматической концентрации ионов Са 2+ , которые продолжают активировать сократительный аппарат и после прекращения электрического процесса.
Что представляет собой сопряжение возбуждения и сокращения (ВС сопряжение)?
Запуск нервным импульсом сокращения скелетной мышцы. При нормальных условиях скелетная мышца в покое слегка натянута. Это свидетельство минимального или слабого связывания актина с миозином. Нервный импульс, достигший терминального нервного окончания, передается на ацетилхолиновый рецептор. В скелетной мышце этот рецептор представлен специализированным образованием, которое называется двигательной концевой пластинкой. Двигательная концевая пластинка представляет собой участок сарколеммы с множеством складок, расположенный в непосредственной близости от нервного окончания. Выделенный нервным окончанием ацетилхолин диффундирует через синаптическую щель и связывается с рецепторами, расположенными на многочисленных складках постсинаптической мембраны (концевой пластинки сарколеммы). Лиганд-рецепторное взаимодействие повышает проницаемость мембраны для натрия, что вызывает местную деполяризацию (потенциал действия концевой пластинки). Потенциал действия концевой пластинки распространяется по сарколемме в разных направлениях и проводится по Т-тру-бочкам внутрь мышечного волокна. Деполяризация триады (концевая цистерна, Т-трубочка и СР) вызывает высвобождение во внутриклеточную жидкость депонированных в СР ионов кальция. При наличии высокой концентрации ионов кальция и достаточного количества энергии запускается цикл поперечных мостиков. Гидролиз вновь синтезированных молекул АТФ реактивирует миозиновые головки, которые присоединяются к другим активным участкам молекулы миозина. Циклическая работа поперечных мостиков продолжается до тех пор, пока имеются свободные ионы кальция и достаточное количество АТФ.
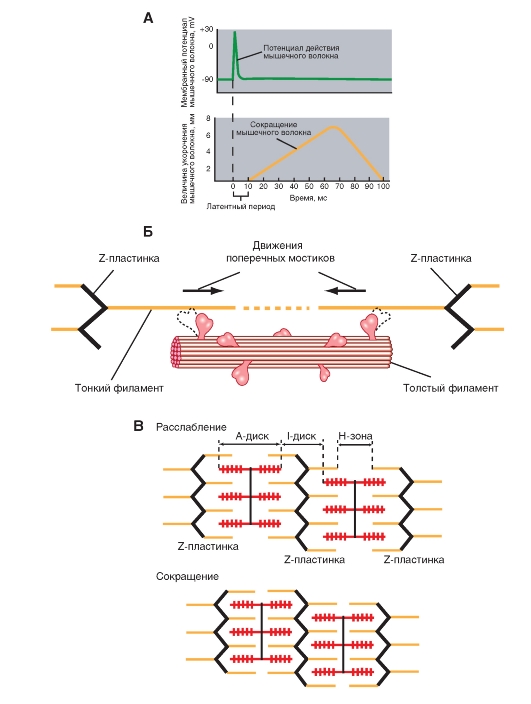
Рис.13. Модель скользящих нитей.
Что такое теория скользящих нитей?
Эта теория объясняет, каким образом фиксированные толстые и тонкие филаменты перемещаются друг относительно друга и обеспечивают сокращение саркомера. Перемещение, происходящее во время цикла поперечных мостиков, обусловлено скольжением молекулы актина по миозину. Повторяющееся присоединение и отделение ряда поперечных мостиков приводит к тому, что параллельно расположенные филаменты скользят друг по другу, сокращая тем самым расстояние между двумя соседними г-линиями. Таким образом саркомер укорачивается. Сокращение саркомера приводит к возникновению некоторой силы.
Модель скользящих нитей
Во время генерирования силы, укорачивающей мышечное волокно, перекрывающиеся толстые и тонкие филаменты каждого саркомера, подтягиваемые движениями поперечных мостиков, сдвигаются друг относительно друга. Длина толстых и тонких филаментов при укорочении саркомера не изменяется (рис. 13). Этот механизм мышечного сокращения известен как модель скользящих нитей.
Механизм электромеханического сопряжения в миокарде. Электрическая активность сердца. Электрокардиограмма.
Механизм электромеханического сопряжения в миокарде. У человека и млекопитающих структуры, которые отвечают за электромеханическое сопряжение в скелетных мышцах, в основном имеются и в волокнах сердца. Для миокарда характерна система поперечных трубочек (Т-система); особенно хорошо она развита в желудочках, где эти трубочки образуют продольные ответвления. Напротив, система продольных трубочек, служащих внутриклеточным резервуаром Са2+, в мышце сердца развита в меньшей степени, чем в скелетных мышцах. Как структурные, так и функциональные особенности миокарда свидетельствуют в пользу тесной взаимосвязи между внутриклеточными депо Са2+ и внеклеточной средой. Ключевым событием в сокращении служит вход в клетку Са2+ во время потенциала действия. Значение этого кальциевого тока состоит не только в том, что он увеличивает длительность потенциала действия и вследствие этого рефрактерного периода: перемещение кальция из наружной среды в клетку создает условия для регуляции силы сокращения. Однако количество кальция, входящего во время ПД, явно недостаточно для прямой активации сократительного аппарата; очевидно, большую роль играет выброс Са2+ из внутриклеточных депо, запускаемый входом Са2+ извне. Кроме того, входящие в клетку ионы пополняет запасы Са2+, обеспечивая последующие сокращения. Таким образом, потенциал действия влияет на сократимость по меньшей мере двумя путями. Он – играет роль пускового механизма («триггерное действие»), вызывающего сокращение путем высвобождения Са2+ (преимущественно из внутриклеточных депо); – обеспечивает пополнение внутриклеточных запасов Са2+ в фазе расслабления, необходимое для последующих сокращений. Механизмы регуляции сокращений. Целый ряд факторов оказывает косвенное влияние на сокращение миокарда, изменяя длительность потенциала действия и тем самым величину входящего тока Са2+. Примеры такого влияния — снижение силы сокращений вследствие укорочения ПД при повышении внеклеточной концентрации К+ или действии ацетилхолина и усиление сокращений в результате удлинения ПД при охлаждении. Увеличение частоты потенциалов действия влияет на сократимость так же, как и повышение их длительности (ритмоинотропная зависимость, усиление сокращений при нанесении парных стимулов, постэкстрасистолическаяпотенциация). Так называемый феномен лестницы (нарастание силы сокращений при их возобновлении после временной остановки) также связан с увеличением внутриклеточной фракции Са2+. Учитывая эти особенности сердечной мышцы, не приходится удивляться тому, что сила сокращений сердца быстро изменяется при изменении содержания Са2+ во внеклеточной жидкости. Удаление Са2+из внешней среды приводит к полному разобщению электромеханического сопряжения; потенциал действия при этом остается почти неизменным, но сокращений не происходит. Ряд веществ, блокирующих вход Са2+ во время потенциала действия, оказывает такой же эффект, как и удаление кальция из внешней среды. К таким веществам относятся так называемые антагонисты кальция (верапамил, нифедипин, дилтиазем) Напротив, при повышении внеклеточной концентрации Са2+ или при действии веществ, увеличивающих вход этого иона во время потенциала действия (адреналин, норадреналин), сократимость сердца увеличивается. В клинике для усиления сердечных сокращений используют так называемые сердечные гликозиды (препараты наперстянки, строфанта и т. д.). В соответствии с современными представлениями сердечные гликозиды повышают силу сокращений миокарда преимущественно путем подавления Nа+/К+-АТФазы (натриевого насоса), что приводит к повышению внутриклеточной концентрации Nа +. В результате снижается интенсивность обмена внутриклеточного Са2+ на внеклеточный Nа+, зависящего от трансмембранного градиента Nа, и Са2+ накапливается в клетке. Это дополнительное количество Са2+ запасается в депо и может быть использовано для активации сократительного аппарата.
Электрокардиография – метод регистрации электрической активности миокарда в ходе сердечного сокращения.
Регистрируется некоторый суммарный эффект активности клеток. На уровне отдельной клетки, при ее переходе в возбужденное состояние, на мембране происходит изменение электрического потенциала: на смену отрицательному потенциалу покоя приходит положительный потенциал действия. Этот процесс сопряжен с переносом ионов различного вида через мембрану. Клеточная мембрана в спокойном состоянии клетки поляризована таким образом: внутри клетки – минус, снаружи – плюс. При переходе клетки в возбужденное состояние ситуация на мембране меняется на противоположную: внутри клетки – плюс (преобладают положительные ионы), снаружи – минус. Эти изменения на клеточной мембране называются деполяризацией.
После того, как возбужденная клетка выполнила свою природную функцию, она возвращается в спокойное состояние; восстанавливаются и начальные концентрации ионов по обе стороны мембраны. Этот процесс называется реполяризацией.
Переходы клеток в возбужденное состояние и последующий их возврат в спокойное состояние носит массовый характер, и в различных участках миокарда начинается, происходит и заканчивается не одновременно. Поэтому можно говорить о волнах деполяризации и реполяризации, распространяющихся по сердцу в целом или по отдельным его частям – предсердиям, желудочкам, перегородкам.
Представлениям о сердце как органе, по которому распространяются волны деполяризации и реполяризации, хорошо соответствует модель, согласно которой сердце – это электрический диполь, электрическое поле которого ощутимо (благодаря проводящим тканям) далеко за пределами сердца, и может регистрироваться с помощью системы электродов, помещенных на поверхности тела.
Источники:
http://studwood.ru/1625505/informatika/elektromehanicheskoe_sopryazhenie_myshtsah
http://helpiks.org/7-65284.html
http://studopedia.ru/19_409893_mehanizm-elektromehanicheskogo-sopryazheniya-v-miokarde-elektricheskaya-aktivnost-serdtsa-elektrokardiogramma.html

